雌激素受体-α 在调节软骨细胞表型和机械负荷反应中的新作用
虽然骨关节炎(OA)的发病机制是多因素的,但关节表面的机械超负荷是 OA 发病和进展的已知驱动因素。软骨细胞衰老被认为是 OA 软骨的一个重要特征。研究表明,人软骨细胞在动态压缩负荷大于 20% 时,可诱导衰老相关标志物的表达,然而,机械负荷影响软骨细胞衰老的机制仍有待阐明。
研究报道,与健康对照组相比,OA 患者软骨细胞上的雌激素受体-α(ERα)水平降低。此外,在 OA 的体外模型中,用促炎细胞因子白细胞介素1β(IL-1β)刺激软骨细胞上调 microRNA 203(miR-203)表达,从而拮抗 ERα 功能,导致炎症增加,细胞活性和软骨生成基质蛋白表达降低。这支持 ERα 在介导软骨细胞表型中的作用。有趣的是,发现静水压力形式的机械负荷可以通过 ERα 调节成骨,这支持通过 ERα 探索软骨形成表型的机械转导信号。
鉴于此,美国匹兹堡大学医学院骨科及中国中南大学湘雅医院骨科的研究团队曾证明了ERα 是 OA 中软骨细胞表型的重要调节因子,包括调节衰老和降解酶的产生。特别是,ERα 缺失的软骨细胞通过显著提高与肥大和成骨相关的分子的表达水平来响应损伤性的机械负荷,这是 OA 软骨中发现的关键特征。因此,恢复和维持软骨细胞中的 ERα 功能可能代表了 OA 的未来治疗干预。研究成果发表在Osteoarthritis and Cartilage期刊题为“Novel role of estrogen receptor-α on regulating chondrocyte phenotype and response to mechanical loading”。

首先,研究人员从接受 TKA(全膝关节置换术)的患者中收集人类 OA 膝关节软骨组织,所有保存的软骨均有轻微至轻度的宏观损伤,对照软骨样本表面完整,受损软骨样本表现出严重的退化。
与对照软骨相比,受损软骨含有较少的糖胺聚糖(GAG),并显示出更高水平的 MMP13、p16INK4a和 SA-β-gal。为了证实组织学结果,从保存的(P-CHs)和受损(D-CHs)软骨中分离出软骨细胞(CHs),进行 qRT-PCR 和蛋白质印迹检测,发现 D-CHs 在衰老、炎症、纤维化、成骨和降解相关基因方面的表达水平高于 P-CHs。蛋白质印迹分析进一步证实了 D-CHs 的衰老和骨关节炎特征,如较高水平的 p16INK4a、p21 和 MMP13。通过检测 P-CHs 和 D-CHs 的软骨形成能力,发现 D-CHs 产生的软骨颗粒通常遗传了受损软骨的表型,如低 GAG 沉积和高水平的细胞衰老(图1 A)。
总的来说,与保存的对照软骨和 P-CHs 衍生组织相比,受损的软骨和 D-CHs 衍生组织表现出更高水平的衰老、炎症、纤维化、成骨和降解。
由于p16INK4a在 D-CHs 中显著上调,因此测试了抑制 p16INK4a是否可以逆转 D-CHs 中的 OA 表型(图1 B)。结果表明,在 2D 软骨细胞培养上,sip16INK4a处理显著降低 p16INK4a水平(图1 F),并部分逆转衰老水平(图1 C、D)。但是,p16INK4a 抑制的 D-CH 颗粒在软骨生成刺激下无法产生软骨(图1 E),尽管软骨生成基因水平没有显著受损(图1 D)。此外,抑制 p16INK4a促进了 MMP13 的表达(图1 D、F)。这说明,在 D-CHs 中抑制 p16INK4a不会增强 D-CHs 在体外生成健康软骨的能力。
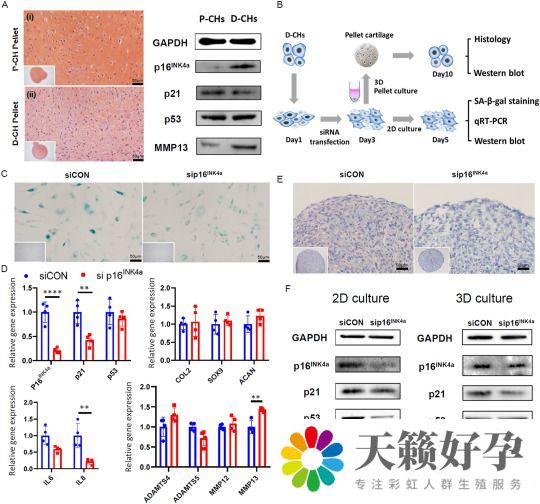
图1 通过siRNA抑制D-CHs中的p16INK4a在逆转D-CHs的不良表型方面
接下来,为了确定对照和受损组软骨之间的差异因子,进行了 RNA-Seq 分析以检查 P-CHs 和 D-CHs 的转录组。结果显示,D-CHs 中软骨降解相关基因的变化最大。有趣的是,雌激素受体家族和雌激素受体-1(ESR1),一种编码雌激素受体-α(ERα)的基因,都位列上游调控因子的前5名。与 P-CHs 相比,D-CHs 的 ESR1 表达显著降低。ERα 的下游靶点显示,许多与软骨功能和 OA 相关的基因被预测受ERα调控,包括p16INK4a。通过检测不同样本中的 ERα 水平,验证了ERα 在严重损伤的软骨中下调。
由于 D-CHs 表现出低 ERα 表达,因此测试了升高 ERα 是否可以逆转 D-CHs 的 OA 表型。结果表明,增强 ERα 水平不仅降低了衰老水平且增加了增殖潜力,同时也改善了 OA 表型。这说明,ERα 可调节软骨细胞表型。
由于软骨损伤最常见于体内机械应力最高的膝关节区域,因此实验还研究了降低的 ERα 水平是否与机械负荷有关,以及降低的 ERα 水平是否改变了软骨细胞对机械负荷的反应。实验过程中使用了0.2Hz,0%、5% 和 20% 的应变,分别代表了静态、中等负荷和损伤性负荷的情况。结果显示,5% 或 20% 动态压缩负荷下,软骨细胞 ESR1 的表达降低(图2 B、C、F)。此外,20% 负荷会导致 siCON 细胞中分解代谢基因(如 ADAMTS4 和 MMP13)的上调,以及衰老标志物(包括p16INK4a和p53)的增加(图2 B、D-F)。这说明,ERα 水平受机械负荷的调节。
考虑到 ERα 对软骨细胞衰老和软骨形成表型的影响,以及其对机械负荷的反应,实验还研究了 ERα 在调节压缩负荷对软骨细胞表型的影响中的作用。结果显示,当存在 ESR1 表达时,20% 应变下的机械负荷不再引起先前看到的 p16INK4a和 MMP13 表达的增加。相反,敲低 ESR1 表达时施加机械负荷会降低这两种分子的表达(图2 B、D-F)。值得注意的是,机械负荷和 ESR1 敲低都增加了 P-CHs 中磷酸化的 p65(pho-p65)的水平,这是细胞应激的代表性标志物。这些数据说明,抑制 ERα 会改变软骨细胞对压缩负荷的反应性。
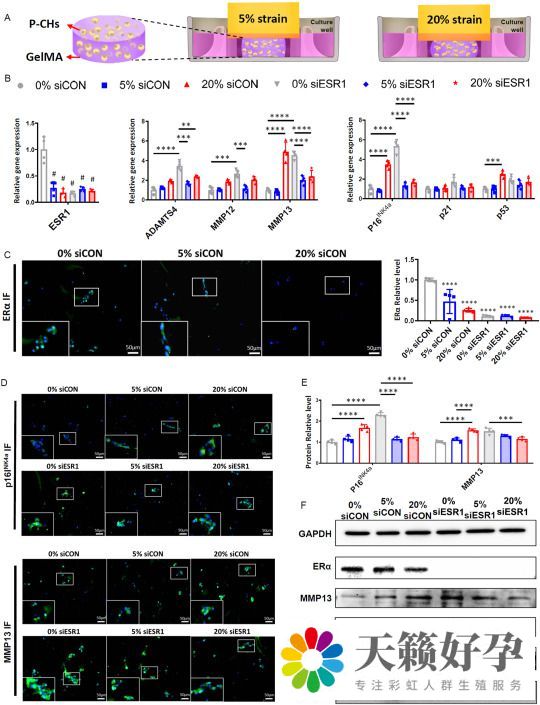
图2 ERα在介导软骨细胞对机械负荷反应中的作用—检测分解代谢
最后,实验进一步研究了机械负荷和 ERα 表达在调节软骨生成、纤维化和成骨基因中的相互作用。当 ESR1 表达保留(siCON)时,5% 和 20% 应变下的机械负荷对成骨或纤维化标记基因的表达没有显著影响,尽管 OSX 和 VCAN 在 20% 压缩应变下显著上调(图3 A)。然而,COL2A1 存在应变反应性的下调(图3 D)。有趣的是,单独敲除 ESR1 (0% siESR1)显著上调了 OCN 的表达,同时抑制了 COL2A1 和软骨寡聚基质蛋白(COMP) 的水平(图3 A-D)。虽然 5% 的应变压缩负荷对 ESR1 敲除没有缓解作用,但20% 的应变倾向于进一步上调肥大相关标志物,包括 COL10 和 OCN(图3 B-D),同时还逆转了 ESR1 敲除对 COL2A1 和 COMP 水平的抑制作用(图3 D)。值得注意的是,与 P-CHs 相比,在人类的 D-CHs 中也观察到 OCN 水平的提高(图3 E、F),并且与野生型对照相比,在从 ESR1 敲除小鼠中分离的软骨中也观察到 OCN 水平增强(图3 E、G)。这说明,ERα 的缺失和压缩超负荷协同增加软骨细胞的肥大转化。
应该注意的是,瞬时受体电位阳离子通道亚家族v 成员4(TRPV4)的表达是一种公认的机械敏感 Ca2+ 通道,对机械负荷也有反应。在 siCON 组中,5% 的负荷显著上调了 TRPV4 的表达,然而,较高的压缩应变(20%)适度降低了 TRPV4 的表达(图3 D)。此外,ESR1 敲除不会改变 TRPV4 蛋白水平,但确实调节了机械负荷的影响,因为增加应变幅度会诱导 TRPV4 表达可观察到的剂量依赖性增加(图3 D)。
基于上述研究的结果,ERα 在调节软骨细胞表型和对机械超负荷反应中的途径如图4所示。
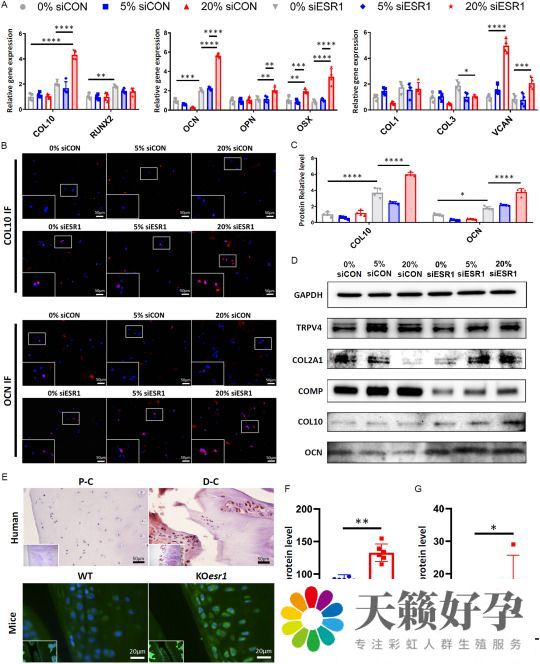
图3 ERα在介导软骨细胞对压缩负荷反应中的作用—检测软骨形成
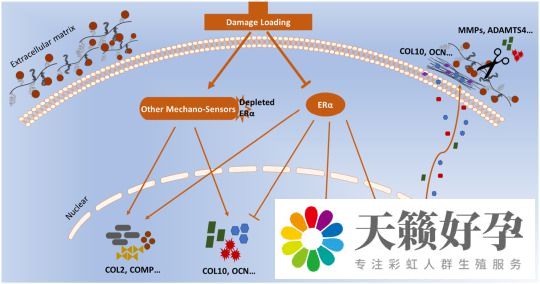
图4 ERα信号通路在调节软骨细胞表型中的作用。
总之,该研究表明,ERα 在介导软骨细胞表型及其对机械负荷的反应中具有新的雌激素非依赖性作用,并表明提高 ERα 水平可能代表了一种治疗骨关节炎的新方法。




